
Researchers from UC Davis collect acorns in arid west Texas to plant on their campus in northern California. They estimate their climate in 2100 will be similar to that of Barstow or even Phoenix today. City staff from a town near Portland, Oregon travel to California and Arizona for seedlings they can take home and plant along their city streets. They are preparing for Portland’s weather to become like Sacramento today.

With these regions breaking new heat records annually – Sacramento just topped 90 degrees for the 110th day (and counting) in 2020—and given that trees take decades to mature, the race is on. Birds can fly, mammals can walk, but trees expand their ranges very slowly. Most acorns from an oak end up within a few hundred yards from their home tree.
Climate velocity, the speed at which ecotones are shifting north, is much faster than that. Our climate is changing ten to one hundred times faster than during a global warming event 55 million years ago known as the Paleocene-Eocene Thermal Maximum (PETM). During that “rapid” spike, palm trees successfully migrated to the Arctic circle, but they had thousands of years to make it there.

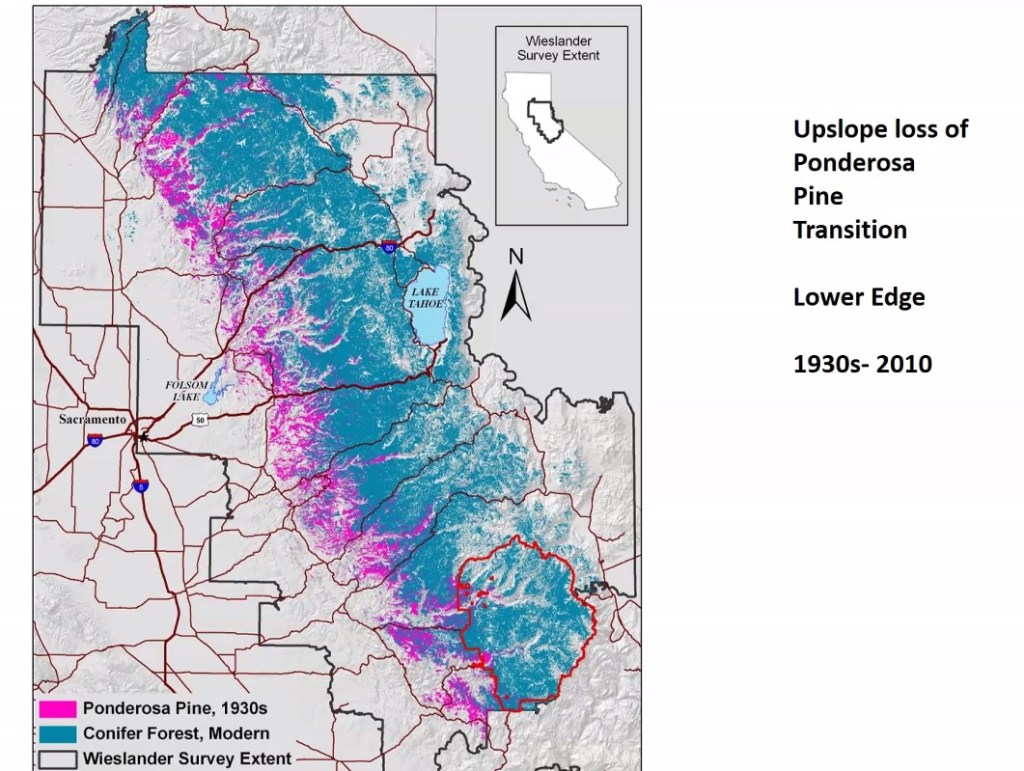
While trees can’t walk, they can die. Range contraction of trees along their southern xeric (dry) edge is happening in the American West right with the speed of climate change. Blue oak die-offs are widespread in the southern third of their range. From California to Colorado, conifers such as Ponderosa pine and Douglas-fir are disappearing from lower elevations. To quote Davis et al (2019), “In areas that have crossed climatic thresholds for regeneration, stand-replacing fires may result in abrupt ecosystem transitions to nonforest states.” When people talk about California becoming Arizona, the cleanup hitter in that process may be fire, but the first batters are heat, drought stress, and bark beetles. After fires, decreased soil moisture and increased vapor pressure deficit (VPD) associated with climate change are leading to reduced probability of regeneration (Davis et al 2019). In short, many forests are not coming back.
Range expansion of trees northward has been documented, but the pace is anemic, insufficient to keep up with the changing climate. One study in the east found that ranges in adult trees expanded north less than 150 yards per year (Sittaro et al 2017). They concluded, “our results add to the body of evidence suggesting tree species are mostly limited in their capacity to track climate warming…”

Researchers have discussed facilitating tree migration due to climate change for over a decade (Aitken et al 2008). For over a hundred years, botanists have recognized regional differences within the same plant species, and simple garden experiments have shown that local varieties do better. The standard rule of thumb has always been that local varieties are best; they are adapted to the local ecological niche. Now that is changing.
Recent research is showing that trees are now in the wrong places; the climate has shifted past them. Valley oaks, white fir, Douglas fir, ponderosa pine, Western hemlock, and lodgepole pine seedlings all do better when removed from their original home and moved north (Aitken and Bemmels 2015).
The local trees are becoming misfits in a world that is changing around them. Many researchers are hesitant to fully embrace assisted migration; introducing non-native species has a horrid track record. But they are beginning to study “assisted gene flow”, moving hardy trees from the southern end of a species’ range to the north end. Cities, on the other hand, are beginning to see trees as more than just aesthetically pleasing; they are critical infrastructure, providing shade and reducing urban temperatures. So the cities and towns are moving faster, boldly cultivating trees from the dry Southwest into the Pacific Northwest. Likewise, the government of British Columbia is not hesitating. Assisted migration of Western larch and whitebark pine in Canada is already underway.

Tree migration is also critical for the range expansion of animals. Without the trees and other vegetation, many birds, mammals, and other forms of life have no habitat rungs on the ladder to enable them to move north as well. Anna’s Hummingbirds now winter in Canada and even Alaska, largely due to ornamental plantings. The Oak Titmouse, on the other hand, is dependent on oaks, tightly constraining its ability to expand north. It may be that, in the coming decades, oaks and other tree species planted in cities and towns will provide critical refugia for a wide variety of birds and insects seeking cooler climes.
 In 2006, I wrote
In 2006, I wrote 
 Many birders suspect this is due to the Founder Effect, a phenomenon that occurs when a small population colonizes a large area. Eventually, all of the birds (or other animal species) are descended from few individuals. In this context, certain recessive traits that were once rare may become more common.
Many birders suspect this is due to the Founder Effect, a phenomenon that occurs when a small population colonizes a large area. Eventually, all of the birds (or other animal species) are descended from few individuals. In this context, certain recessive traits that were once rare may become more common.
